【 Abstract 】 To investigate the effects of excessive reduced Glutathione (GSH) on Lung fibroblasts (LFS), in order to explore the status and changes of LFS after the REDOX balance is broken.
Fresh rat LFS were extracted, and the effects of different concentrations of GSH on the proliferation, growth, migration and signaling pathway of LFS were observed by cell viability assay, gelatin enzyme assay, cell morphology assay, scratch assay, cell immunofluorescence assay and protein immunoimprinting assay.
The results show that extracellular GSH intervention can induce changes in the homeostasis of LFS, enabling local adhesion kinase (FAK), Nuclear factor kappa-B, adhesion and adhesion of LFS to Focal adhesion kinase (FAK). The expression of NF-κB and Phospho-histone gamma H2A.X, γ-H2A.X increased, but inhibited the activity of matrix metalloproteinase-2/9 (MMP-2/9) secreted by cells. The cell migration, the expression of mitogen-activated protein kinase (p38) and the expression of tumor suppressor gene (p53) protein were inhibited.
Excessive extracellular GSH may lead to DNA damage and cell failure and dysfunction of LFS, resulting in lung tissue lesions.
The use of too many antioxidants will disrupt the homeostasis of cells and may cause irreversible damage to the body.
【 Key words 】 Reduced glutathione; Pulmonary fibrosis; Lung fibroblasts; REDOX equilibrium
Glutathione highlights
1) High concentration of glutathione caused DNA damage and growth arrest in lung fibroblasts.
2) High concentration of glutathione can inhibit the activity of matrix metalloproteinase-2/9 (MMP-2/9), leading to the disorder of extracellular matrix.
3) Glutathione is commonly used as an antioxidant in pharmaceutical foods, and overuse can cause cell and tissue damage.
Research background
Pulmonary fibrosis (PF) is a chronic, irreversible, and fatal disease caused by various lung diseases or other diseases, which refers to the process of alveolar damage, excessive repair of lung tissue, and abnormal remodeling.

These structural changes accelerate fibrosis in the surrounding area, causing lung damage and abnormal lung function, which is ultimately life-threatening.
Glutathione plays an important role in cell differentiation, proliferation and apoptosis, and is the main regulator of cell REDOX state and REDOX signal transduction.
REDOX disorders in cells associated with many diseases, such as inflammation, immunity, metabolism, etc. The occurrence of oxidative stress reduces the metabolism of Glutathione (GSH), and the reduced ratio of reduced and oxidized glutathione leads to mitochondrial dysfunction, which is involved in the regulation of iron metabolism and REDOX imbalance in iron death.
Glutathione concentration too high or too low will cause the REDOX balance to disrupted, resulting in the body’s ability to respond to oxidative stress reduced.
Studies have found that GSH in lung tissue of patients with pulmonary fibrosis elevated and the oxidative metabolic balance broken, but the effect of increased GSH concentration on cells and the mechanism of influence are still unclear.
Lung fibroblasts (LFS) are key cells in the process of PF and the construction and maintenance of Extracellularmatrix (ECM). Lung abnormalities cause LFS to undergo abnormal differentiation, transformation and proliferation, and secrete collagen and other ECM components in large quantities. Leading to ECM deposition and remodeling, thus impairs lung function, which is a key event in the PF process and an important reason for the progression of PF.
In order to investigate the effect of extracellular glutathione on LFS and its role in the development and development of PF, a cell model established using freshly extracted rat LFS.
Key content
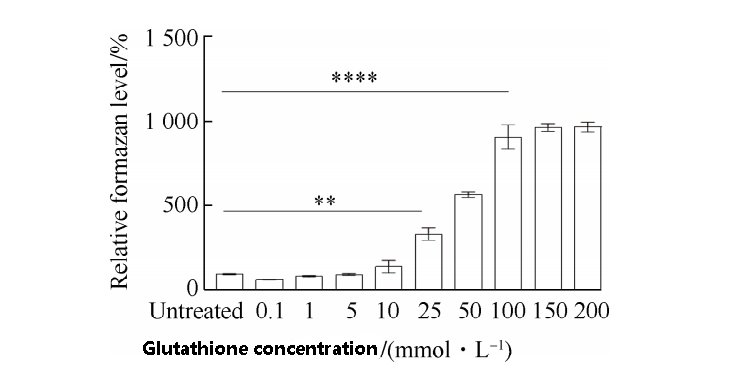
Compared with the untreated cells, a gradual increase in glutathione concentration increases the cytoskeletal level,
indicating that glutathione enhances the activity of intracellular dehydrogenase and interferes with cellular REDOX homeostasis.
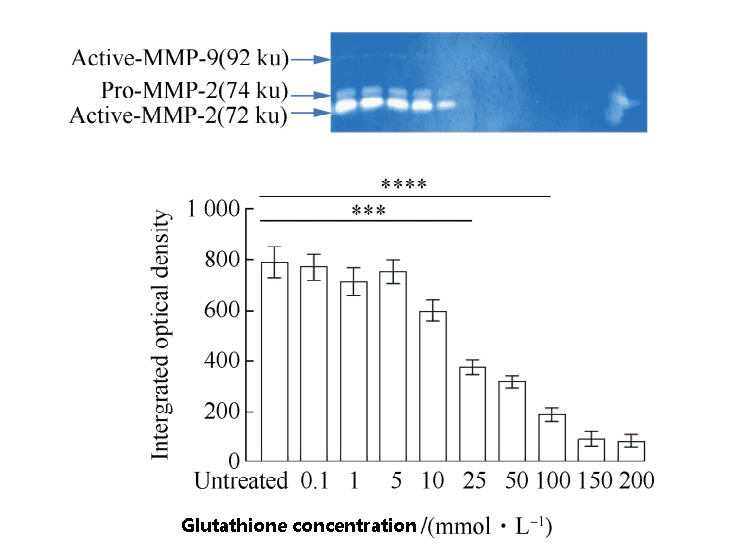
The inhibitory effects of different concentrations of GSH on matrix metalloproteinases in lung fibroblasts shown in Figure 2.
When the concentration of GSH low, the activity of MMP-2/9 basically unchanged, and when the concentration of GSH more than 10 mmol/L,
the activity of MMP-2/9 significantly decreased, indicating that the degradation of extracellular matrix affected, and its morphology also affected.
The morphological results shown in the figure.
With the increase of time, when the concentration of glutathione was 1 mmol/L, there was no significant change in cell morphology.
When the concentration of glutathione was 10 mmol/L, the cell morphology showed different degrees of contraction,
the gap between cells gradually increased, the number of cells decreased, and the cell death occurred after 4 h.
However, when the concentration of glutathione was 100 mmol/L,
the cells had no significant change, no increase or decrease, and the cells were in a stagnant state.

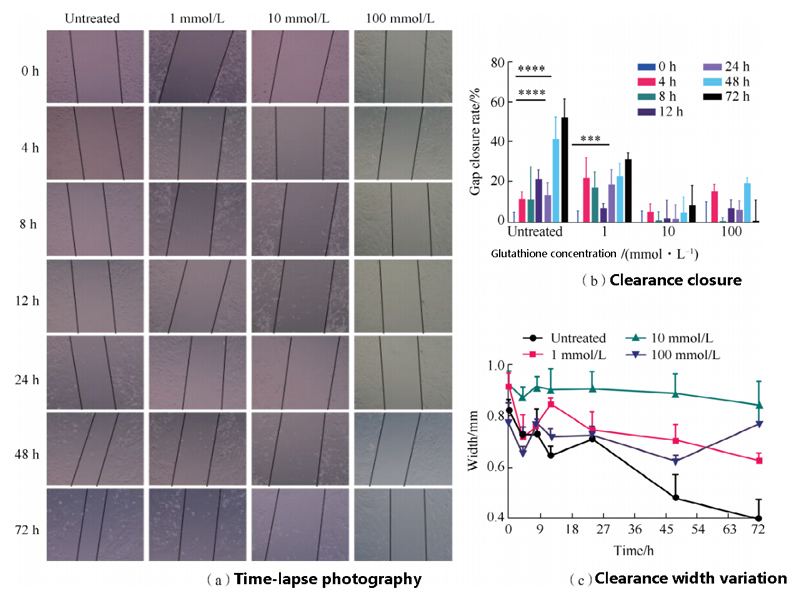
In order to evaluate the effects of different concentrations of GSH on LFS migration,
time-lapse photography used to detect cell morphology for 72 h, and the results shown in the figure.
The results showed that the cell space of the control group decreased gradually with the increase of time.
When GSH concentration was 1 mmol/L, the cell space of the control group decreased with the increase of time,
but the degree of reduction was smaller than that of the control group.
When the concentration of GSH was 10mmol/L,
the cell gap did not change significantly with the increase of time, and the cells showed atrophy.
When glutathione concentration 100 mmol/L, the cell gap width basically unchanged,
the cell gap closure rate basically unchanged, and the cells were almost in a stagnant state.
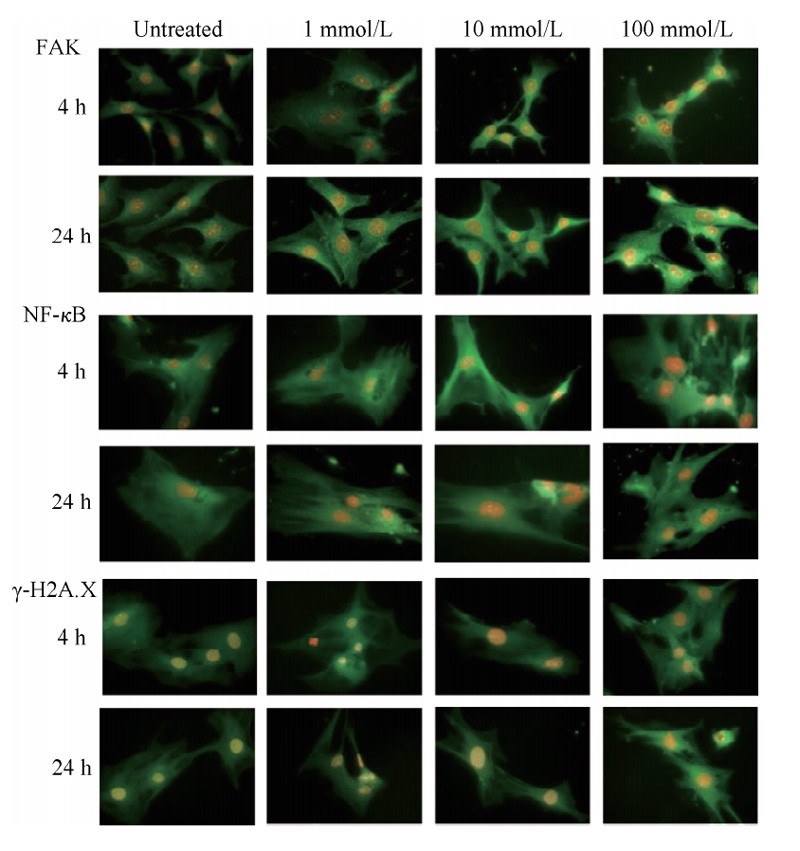
The effect of GSH on the expression of signaling protein in LFS investigated by immunofluorescence staining and protein imprinting analysis.
As shown in Figure 5, GSH induced Focal adhesion kinase (FAK), Nuclear factor kappa-B (NF-κB),
focal adhesion kinase (FAK) and nuclear factor Kappa-B (NF-κB). The expression of Phospho-histone gamma H2A.X, γ-H2A.X increased and the fluorescence intensity increased at 24 h.
These results suggest that high extracellular GSH concentration may lead to enhanced migration and adhesion of LFS,
accelerated synthesis of proinflammatory factors and DNA damage.
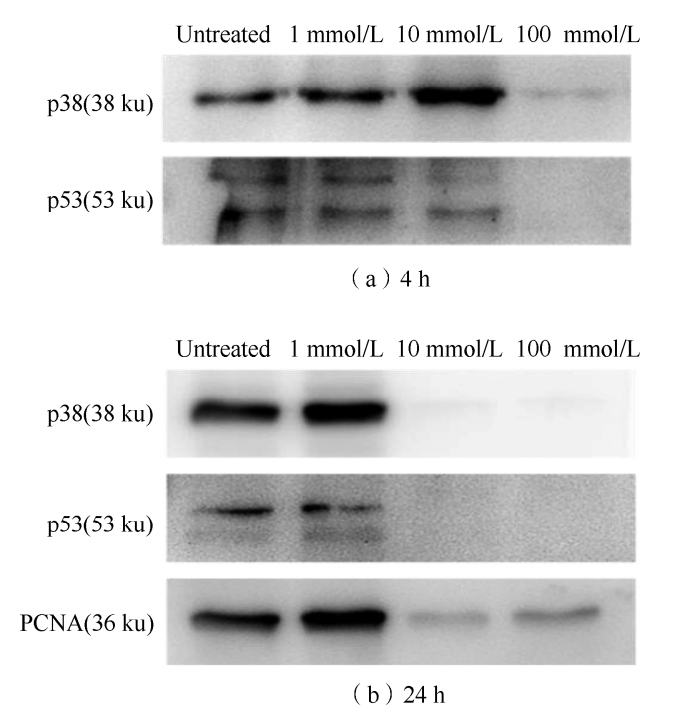
Protein immunoimprinting showed that when GSH treated for 4 h,
the expression of p38 gradually increased with the increase of GSH concentration,
but when GSH concentration reached 100 mmol/L, the expression of p38 disappeared.
With the increase of GSH concentration, the expression of p53 decreased,
and when the GSH concentration was 100 mmol/L, the expression of p53 also disappeared.
When GSH treated for 24 h, the expressions of p38 and PCNA increased when GSH concentration 1 mmol/L,
but when GSH concentration 10,100 mmol/L, the expressions of p38 and PCNA almost non-detected.
The expression of p53 decreased at GSH concentration of 1 mmol/L,
while almost no expression of p53 found at GSH concentration of 10,100 mmol/L.
These may mean that cell stasis or loss of function may occur when extracellular GSH concentrations are high.
Under normal conditions, cell tissues and the body maintain homeostasis, and if homeostasis changed, lesions will occur. The increase of GSH level in lung tissue will affect the activation and differentiation of LFS, leading to pulmonary fibrosis.
conclusion
The occurrence and development of pulmonary fibrosis may be related to the increase of glutathione level in lung tissue and its effect on LFS.
In addition, glutathione, as a commonly used antioxidant, can protect cells from the damage of reducing agents,
but excessive antioxidant use can also cause cell damage and cause unpredictable consequences for the body.